
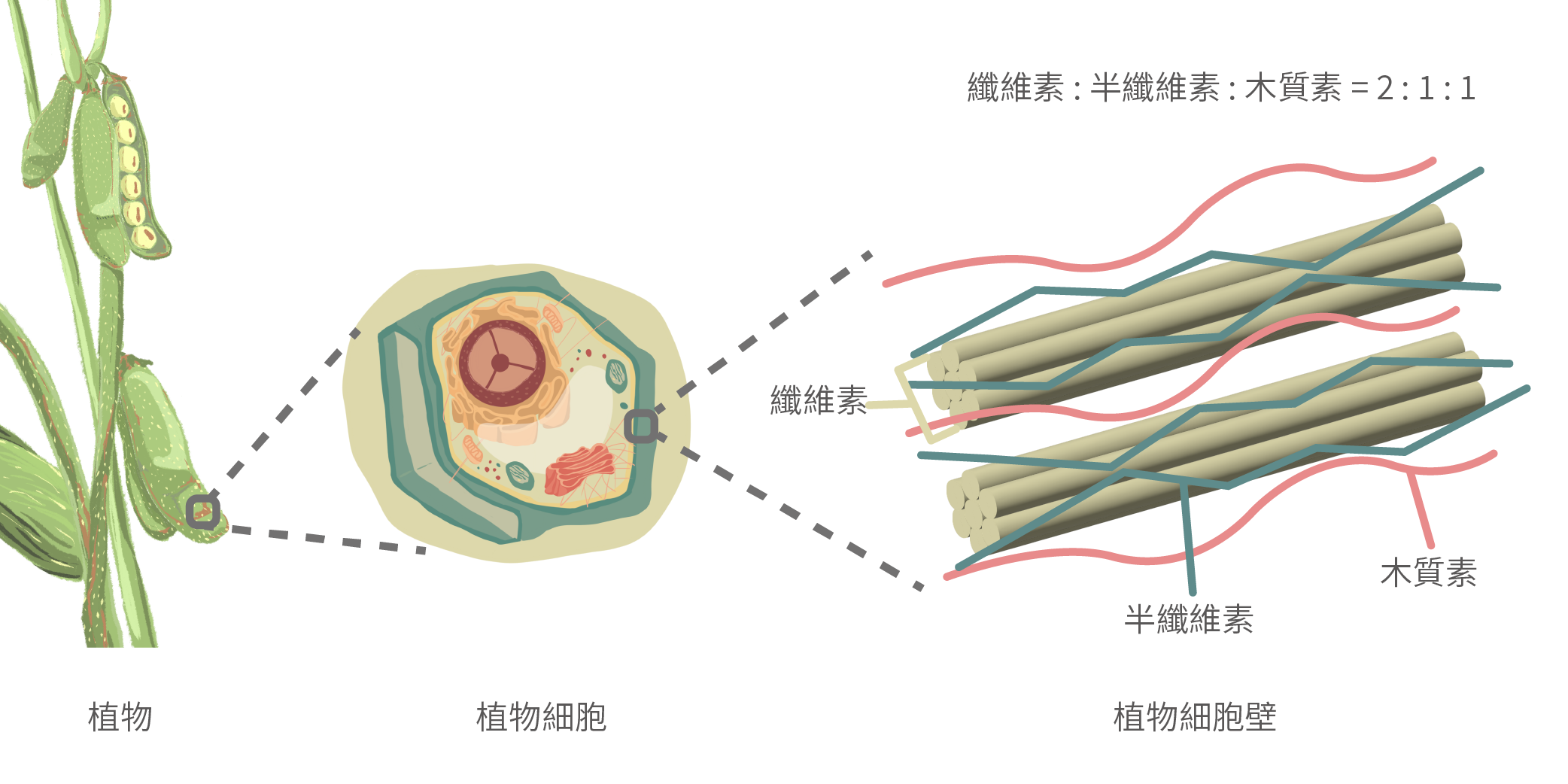
結 構性植物材料由纖維素、半纖維素和木質素構成,其比例大約為2:1:1。
纖維素以不溶性束狀的多醣鏈存在,提供細胞壁的強度和剛性,而木質素則作為富含芳香化合物的無定形屏障,阻礙微生物的降解。半纖維素與纖維素和木質素交織在一起,形成一個矩陣,為細胞壁提供抵抗微生物降解的能力,同時保持足夠的靈活性,以允許細胞移動和生長(Van Zyl等,2010)。半纖維素是由D-木糖、D-半乳糖、D-甘露糖、D-葡萄糖和L-阿拉伯糖組成的線性和分支多醣。根據聚合物骨架中主要存在的單醣,半纖維素被分類為木聚糖、半乳聚糖和甘露聚糖(Gray等,2006)。作為一種半纖維素多醣,甘露聚糖在豐富度上僅次於木聚糖(McCleary, 1988)。甘露聚糖家族包括四個亞家族:葡甘露聚糖、半乳甘露聚糖、半乳葡甘露聚糖和純甘露聚糖(Petkowicz等,2001)。甘露聚糖和異甘露聚糖是所有豆科植物細胞壁半纖維素部分的一部分(Reid, 1985),包括豆粕(SBM)、芝麻粕、棕櫚仁粕、椰子粕和瓜爾粕(Dhawan和Kaur, 2007)。豆粕中的β-甘露聚糖是由重複的β-1,4-甘露糖和與β-甘露聚糖骨架相連的α-1,6-半乳糖分子形成的線性多醣(Jackson等,2004)。
飼料中的非澱粉多醣(NSP)對家禽來說是不可消化的,但通過添加酶可以將其作為潛在的能量來源(Meng等,2005)。除了作為潛在的能量來源外,飼料中的NSP,因為單胃動物無法消化,會導致腸道黏度增加、營養消化率降低、飼料轉換率(FCR)增加,最終導致動物表現下降(Bedford和Classen, 1992;Bedford和Morgan, 1996;Lázaro等,2003)。
豆粕是主要的植物蛋白來源,含有3%可溶性NSP和16%不可溶性NSP(Irish和Balnave, 1993),其主要成分是甘露聚糖和半乳甘露聚糖(Slominski, 2011)。β-甘露聚糖或半乳甘露聚糖是多醣,由含有半乳糖的重複甘露糖單位組成。 豆粕中的半乳甘露聚糖含量已報告在1.02%至1.51%之間(Hsiao等,2006)。雖然豆粕中的半乳甘露聚糖含量較低,但由於其具有抗營養性質,仍然引起營養學家的關注。甘露聚糖是多種病原體的表面成分,先天免疫系統會對這些病原體上的抗原作出反應。飼料中的甘露聚糖可能會刺激先天免疫系統,導致無效且耗能的免疫反應。

甘露聚糖的主要成分是D-甘露糖,這是一種六碳糖。由於植物甘露聚糖的異質性和複雜的化學結構,單胃動物無法將其分解成簡單糖作為能量來源,這需要內切1,4-β-甘露聚糖酶、外切β-甘露苷酶(Dhawan和Kaur, 2007)和β-葡萄糖苷酶(Moreira和Filho, 2008)的協同作用。內切β-甘露聚糖酶在自然界中廣泛存在,是一種內水解酶,通過切割甘露聚糖骨架內部的糖苷鍵,產生β-1,4-甘露寡糖(Chauhan等,2012)。水解程度受多種因素影響,包括甘露聚糖骨架中半乳糖的取代程度和分佈(Stalbrand等,1993)、乙酰化程度(McCleary, 1991)、β-甘露聚糖酶的來源(如細菌、真菌等)(McCleary, 1979)以及酶的數量。對於動物營養,最重要的甘露聚糖和半乳甘露聚糖分解酶是內切β-甘露聚糖酶**和α-半乳糖苷酶,因為在家禽營養中使用的飼料中含有半乳甘露聚糖和/或線性甘露聚糖(Shastak等,2015)。
目前,甘露聚糖酶在工業過程中有許多應用,主要用於提高食品品質、改善飼料營養價值,以及在造紙和紙漿工業中進行酶漂白(Dhawan和Kaur, 2007)。具體應用包括:從木纖維中提取木質素、降低咖啡萃取物的黏度、用於洗滌劑工業中的去污增效劑、提高油氣流動性、提取椰肉中的油脂、分解增稠劑以及提高動物飼料的營養價值。許多研究已發表,探討了β-甘露聚糖酶對家禽表現和飼料消化率的影響(Jackson等,1999,2002a,2004;Odetallah等,2002;Lee等,2003a;Daskiran等,2004;Wu等,2005;Li等,2010);但最近的研究集中在雙酶應用(Williams等,2014),減少飼料引起的免疫反應的可能性(Arsenault等,2017),以及確定最低基質水平(半乳甘露聚糖),以引發對動物的負面影響(Latham等,2018)。
作用機制
要確定β-甘露聚糖酶在單胃動物生產系統中的長期可行性和潛在效益,營養學家必須考慮其作用機制及對生產系統的潛在益處。對於β-甘露聚糖酶,提出並定義了多種作用機制,包括:減少動物消化道中的黏度,抑制有害微生物的增殖,以及降低由植物成分中的半乳甘露聚糖引發的飼料誘導免疫反應。這些機制有助於改善營養消化和利用率,並提高動物的生長性能和效率。
1. 減少黏度:
動物消化道中的高黏度會對營養吸收和動物表現產生負面影響(Smits 等,1997;Lee等,2003a, b;Latham等,2018)。黏度增加會影響消化道內消化物的物理混合能力,阻礙消化酶的擴散和對流運輸,從而減少營養與其相應消化分泌物的接觸強度(Choct和Annison, 1992)。例如,半乳甘露聚糖具有很強的吸水性,會形成高黏度的消化物。研究顯示,飼料中溶解性半乳甘露聚糖的增加會提高黏度,減少葡萄糖吸收,並增加飼料轉換率(FCR)(Latham等,2018)。通過β-甘露聚糖酶的添加,可以有效水解半乳甘露聚糖並降低動物腸道中的黏度,從而提高營養吸收(Shastak等,2015)。
2. 免疫反應的抑制:
某些植物中的甘露聚糖(如豆粕中的半乳甘露聚糖)會引發動物 的先天免疫反應,這會導致能量的消耗,進而影響動物的表現(Dale等,2008)。這些植物源的甘露聚糖被視為病原相關分子模式(PAMPs)的類似物,會激活單核細胞和巨噬細胞,並促使細胞因子的生成。β-甘露聚糖酶的補充可以減少免疫系統的激活,從而提高能量的利用率和動物的生長性能(Wu等,2005;Li等,2010)。近期的研究也證實了,添加β-甘露聚糖酶能減少由飼料誘導的免疫反應(Arsenault等,2017)。
3. 抗病原體:
β-甘露聚糖酶將甘露聚糖水解生成短鏈β-1,4-甘露寡糖和D-甘露 糖,這些水解產物可以通過發酵轉化為宿主的能量來源,或結合到病原細菌上的甘露糖專一性結合位點,防止病原菌在消化道中的定殖(Ofek等,1977)。例如,在一項研究中,β-甘露聚糖酶的添加減少了雞隻對沙門氏菌的定殖(Gutierrez等,2008)。
4. NSP降解功能:
β-甘露聚糖酶還具有典型的非澱粉多醣(NSP)降解功能,能提高 飼料中的營養消化率,類似於木聚糖酶的作用。NSP降低營養消化率的機制之一是“籠效應(cage effect)”,即NSP將澱粉和蛋白質等營養物質包裹在胚乳內,阻礙其消化(Bedford, 1993;Slominski, 2011)。雖然“籠效應”在豆類中的影響較小,但多項研究表明,添加β-甘露聚糖酶可以增加胺基酸和能量的消化率(Ferreira等,2016;Daskiran等,2004)。
資料來源: Enzyme in Farm Animal Nutrition, 3rd Edition
結 構性植物材料由纖維素、半纖維素和木質素構成,其比例大約為2:1:1。
纖維素以不溶性束狀的多醣鏈存在,提供細胞壁的強度和剛性,而木質素則作為富含芳香化合物的無定形屏障,阻礙微生物的降解。半纖維素與纖維素和木質素交織在一起,形成一個矩陣,為細胞壁提供抵抗微生物降解的能力,同時保持足夠的靈活性,以允許細胞移動和生長(Van Zyl等,2010)。半纖維素是由D-木糖、D-半乳糖、D-甘露糖、D-葡萄糖和L-阿拉伯糖組成的線性和分支多醣。根據聚合物骨架中主要存在的單醣,半纖維素被分類為木聚糖、半乳聚糖和甘露聚糖(Gray等,2006)。作為一種半纖維素多醣,甘露聚糖在豐富度上僅次於木聚糖(McCleary, 1988)。甘露聚糖家族包括四個亞家族:葡甘露聚糖、半乳甘露聚糖、半乳葡甘露聚糖和純甘露聚糖(Petkowicz等,2001)。甘露聚糖和異甘露聚糖是所有豆科植物細胞壁半纖維素部分的一部分(Reid, 1985),包括豆粕(SBM)、芝麻粕、棕櫚仁粕、椰子粕和瓜爾粕(Dhawan和Kaur, 2007)。豆粕中的β-甘露聚糖是由重複的β-1,4-甘露糖和與β-甘露聚糖骨架相連的α-1,6-半乳糖分子形成的線性多醣(Jackson等,2004)。
飼料中的非澱粉多醣(NSP)對家禽來說是不可消化的,但通過添加酶可以將其作為潛在的能量來源(Meng等,2005)。除了作為潛在的能量來源外,飼料中的NSP,因為單胃動物無法消化,會導致腸道黏度增加、營養消化率降低、飼料轉換率(FCR)增加,最終導致動物表現下降(Bedford和Classen, 1992;Bedford和Morgan, 1996;Lázaro等,2003)。
豆粕是主要的植物蛋白來源,含有3%可溶性NSP和16%不可溶性NSP(Irish和Balnave, 1993),其主要成分是甘露聚糖和半乳甘露聚糖(Slominski, 2011)。β-甘露聚糖或半乳甘露聚糖是多醣,由含有半乳糖的重複甘露糖單位組成。 豆粕中的半乳甘露聚糖含量已報告在1.02%至1.51%之間(Hsiao等,2006)。雖然豆粕中的半乳甘露聚糖含量較低,但由於其具有抗營養性質,仍然引起營養學家的關注。甘露聚糖是多種病原體的表面成分,先天免疫系統會對這些病原體上的抗原作出反應。飼料中的甘露聚糖可能會刺激先天免疫系統,導致無效且耗能的免疫反應。
甘露聚糖的主要成分是D-甘露糖,這是一種六碳糖。由於植物甘露聚糖的異質性和複雜的化學結構,單胃動物無法將其分解成簡單糖作為能量來源,這需要內切1,4-β-甘露聚糖酶、外切β-甘露苷酶(Dhawan和Kaur, 2007)和β-葡萄糖苷酶(Moreira和Filho, 2008)的協同作用。內切β-甘露聚糖酶在自然界中廣泛存在,是一種內水解酶,通過切割甘露聚糖骨架內部的糖苷鍵,產生β-1,4-甘露寡糖(Chauhan等,2012)。水解程度受多種因素影響,包括甘露聚糖骨架中半乳糖的取代程度和分佈(Stalbrand等,1993)、乙酰化程度(McCleary, 1991)、β-甘露聚糖酶的來源(如細菌、真菌等)(McCleary, 1979)以及酶的數量。對於動物營養,最重要的甘露聚糖和半乳甘露聚糖分解酶是內切β-甘露聚糖酶**和α-半乳糖苷酶,因為在家禽營養中使用的飼料中含有半乳甘露聚糖和/或線性甘露聚糖(Shastak等,2015)。
目前,甘露聚糖酶在工業過程中有許多應用,主要用於提高食品品質、改善飼料營養價值,以及在造紙和紙漿工業中進行酶漂白(Dhawan和Kaur, 2007)。具體應用包括:從木纖維中提取木質素、降低咖啡萃取物的黏度、用於洗滌劑工業中的去污增效劑、提高油氣流動性、提取椰肉中的油脂、分解增稠劑以及提高動物飼料的營養價值。許多研究已發表,探討了β-甘露聚糖酶對家禽表現和飼料消化率的影響(Jackson等,1999,2002a,2004;Odetallah等,2002;Lee等,2003a;Daskiran等,2004;Wu等,2005;Li等,2010);但最近的研究集中在雙酶應用(Williams等,2014),減少飼料引起的免疫反應的可能性(Arsenault等,2017),以及確定最低基質水平(半乳甘露聚糖),以引發對動物的負面影響(Latham等,2018)。
作用機制
要確定β-甘露聚糖酶在單胃動物生產系統中的長期可行性和潛在效益,營養學家必須考慮其作用機制及對生產系統的潛在益處。對於β-甘露聚糖酶,提出並定義了多種作用機制,包括:減少動物消化道中的黏度,抑制有害微生物的增殖,以及降低由植物成分中的半乳甘露聚糖引發的飼料誘導免疫反應。這些機制有助於改善營養消化和利用率,並提高動物的生長性能和效率。
1. 減少黏度:
動物消化道中的高黏度會對營養吸收和動物表現產生負面影響(Smits 等,1997;Lee等,2003a, b;Latham等,2018)。黏度增加會影響消化道內消化物的物理混合能力,阻礙消化酶的擴散和對流運輸,從而減少營養與其相應消化分泌物的接觸強度(Choct和Annison, 1992)。例如,半乳甘露聚糖具有很強的吸水性,會形成高黏度的消化物。研究顯示,飼料中溶解性半乳甘露聚糖的增加會提高黏度,減少葡萄糖吸收,並增加飼料轉換率(FCR)(Latham等,2018)。通過β-甘露聚糖酶的添加,可以有效水解半乳甘露聚糖並降低動物腸道中的黏度,從而提高營養吸收(Shastak等,2015)。
2. 免疫反應的抑制:
某些植物中的甘露聚糖(如豆粕中的半乳甘露聚糖)會引發動物 的先天免疫反應,這會導致能量的消耗,進而影響動物的表現(Dale等,2008)。這些植物源的甘露聚糖被視為病原相關分子模式(PAMPs)的類似物,會激活單核細胞和巨噬細胞,並促使細胞因子的生成。β-甘露聚糖酶的補充可以減少免疫系統的激活,從而提高能量的利用率和動物的生長性能(Wu等,2005;Li等,2010)。近期的研究也證實了,添加β-甘露聚糖酶能減少由飼料誘導的免疫反應(Arsenault等,2017)。
3. 抗病原體:
β-甘露聚糖酶將甘露聚糖水解生成短鏈β-1,4-甘露寡糖和D-甘露 糖,這些水解產物可以通過發酵轉化為宿主的能量來源,或結合到病原細菌上的甘露糖專一性結合位點,防止病原菌在消化道中的定殖(Ofek等,1977)。例如,在一項研究中,β-甘露聚糖酶的添加減少了雞隻對沙門氏菌的定殖(Gutierrez等,2008)。
4. NSP降解功能:
β-甘露聚糖酶還具有典型的非澱粉多醣(NSP)降解功能,能提高 飼料中的營養消化率,類似於木聚糖酶的作用。NSP降低營養消化率的機制之一是“籠效應(cage effect)”,即NSP將澱粉和蛋白質等營養物質包裹在胚乳內,阻礙其消化(Bedford, 1993;Slominski, 2011)。雖然“籠效應”在豆類中的影響較小,但多項研究表明,添加β-甘露聚糖酶可以增加胺基酸和能量的消化率(Ferreira等,2016;Daskiran等,2004)。
資料來源: Enzyme in Farm Animal Nutrition, 3rd Edition